
Artículo dedicado al Dr. D. Adolfo García Sastre, el “Doctor Bacteria” del siglo XXI y su Escuela Científica todos ellos decididos Cajalianos. Adolfo es Socio de Honor de Alumni, Huésped Distinguido de la ciudad de Salamanca entre otros muchos y merecidos honores.
«El placer indescriptible —que empalidece el resto de las alegrías de la vida— es una abundante compensación para el investigador que soporta el doloroso y perseverante trabajo analítico que precede a la aparición de la nueva verdad, como el dolor del parto. Es cierto decir que nada para el erudito científico es comparable a las cosas que ha descubierto»
Santiago Ramón y Cajal
En el año 1903, Santiago Ramón y Cajal publicó en los Trabajos del Laboratorio de Investigaciones Biológicas un artículo que, en apariencia, era puramente técnico: “Un sencillo método de coloración selectiva del retículo protoplasmático”. En él, detallaba con su meticulosidad característica un nuevo método de impregnación con nitrato de plata que prometía resultados constantes y aplicables a todo el sistema nervioso. Sin embargo, entre las descripciones de soluciones y tiempos de fijación, su mirada infatigable, entrenada para discernir el orden en el caos, se detuvo en un detalle minúsculo que había pasado desapercibido para todos. Al describir el núcleo de las neuronas, anotó: “…todas las neuronas, sin excepción, exhiben otro corpúsculo especial esférico, pequeñísimo, generalmente único… Semejantes granitos, que por llamarlos de algún modo los denominaremos cuerpos intranucleares accesorios”.
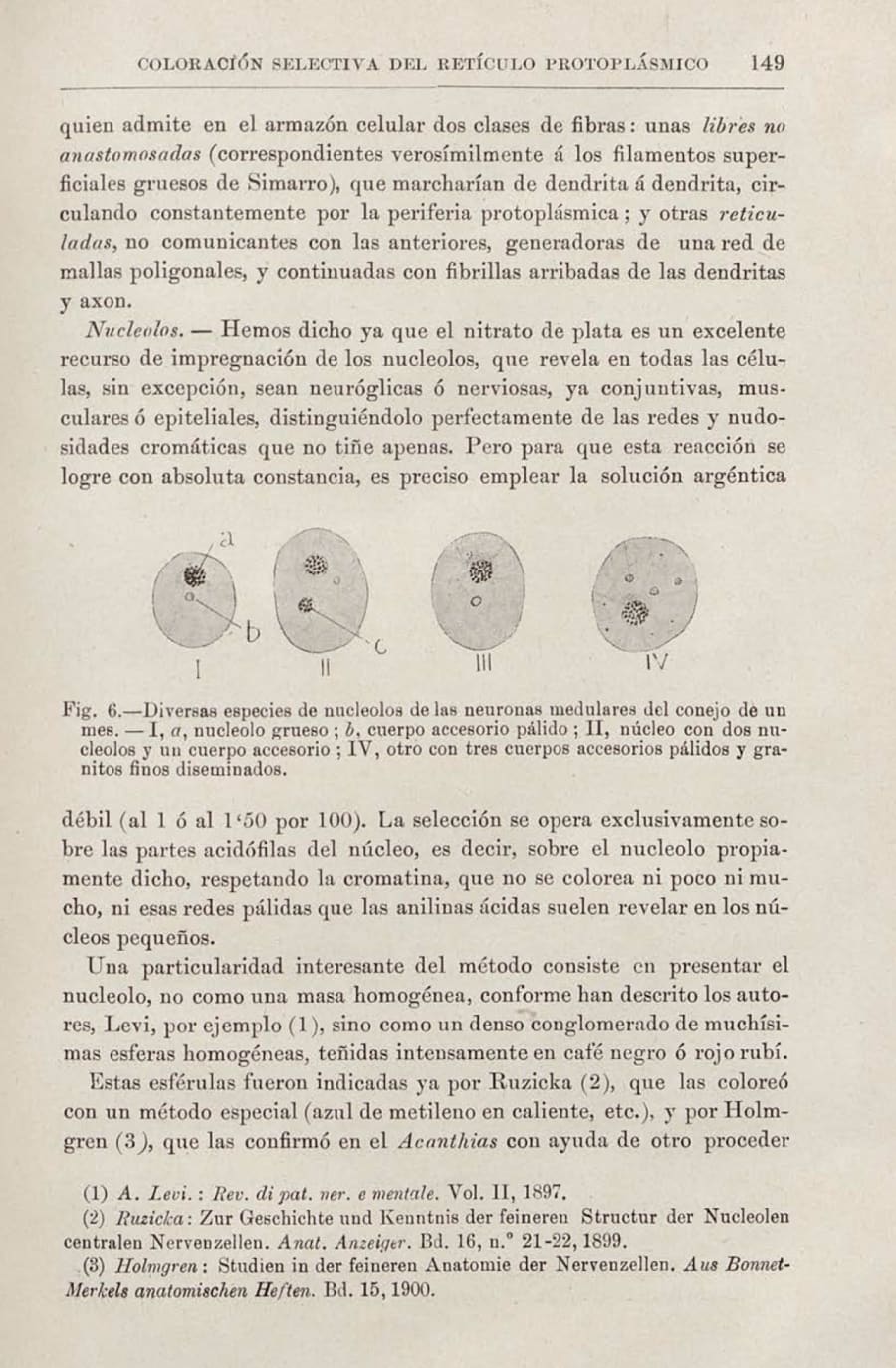


Observó que estas pequeñas esferas se teñían de un tono “amarillo o anaranjado”, distinto al negro del nucleolo, y concluyó que su composición química “no es idéntica”. Este descubrimiento no fue un accidente, sino la consecuencia ineludible de su singular método científico: una fusión sublime de observación empírica rigurosa y una profunda, casi mística, sensibilidad artística.
El Ojo del Artista, la Mente del Científico
Cajal sostenía con fervor que todo neuroanatomista debía ser un artista, pues el acto de dibujar no era un mero registro, sino una herramienta de indagación, una forma de pensamiento visual. A diferencia de la fría e imparcial fotografía, el dibujo le permitía al científico-artista sintetizar múltiples observaciones, interpretar la arquitectura tridimensional y, lo que es más importante, discernir el orden en el caos del «enmarañado y misterioso bosque» del sistema nervioso. Fue precisamente su nuevo método de tinción, detallado en su publicación de 1903 (artículo adjunto al final del artículo), el que le confirió la claridad de visión necesaria para identificar estructuras que habían pasado desapercibidas para otros. Esta «disciplina del ojo», cultivada a través del dibujo, no era una habilidad para registrar, sino para comprender. La capacidad de Cajal para abstraer y construir un modelo mental coherente a partir de datos fragmentarios fue la misma que le permitió ver las neuronas como entidades individuales, derribando la teoría reticular de Golgi, y la que le permitió identificar estos sutiles corpúsculos que otros, con una mirada menos poética, podrían haber descartado como meros artefactos. Su arte no era un subproducto de su ciencia; era la herramienta que la posibilitaba.
Esta perspectiva tiene una profunda resonancia en la ciencia contemporánea. En una era dominada por la “big data” y tecnologías de imagen de resolución sin precedentes, la capacidad humana para interpretar, conceptualizar y encontrar patrones —el “ojo del artista”— sigue siendo insustituible. El legado de Cajal nos enseña que la verdadera comprensión no emerge de la mera acumulación de datos, sino de su interpretación significativa. Más de un siglo después, esa observación, nacida de la confluencia entre el arte y la ciencia, ha florecido hasta convertirse en un campo de una importancia crítica y sorprendente. El «cuerpo accesorio», hoy conocido universalmente como el Cuerpo de Cajal, se revela como un epicentro estratégico, un campo de batalla central en la incesante carrera armamentística entre la célula huésped y los virus invasores. Este artículo explora cómo el legado oculto de Cajal ha transitado desde una curiosidad citológica hasta la vanguardia de la virología y la inmunidad nuclear, trazando una línea directa desde la filosofía del sabio español hasta los laboratorios que hoy combaten las pandemias del futuro.
Un Condensado de Vida en el Corazón del Núcleo
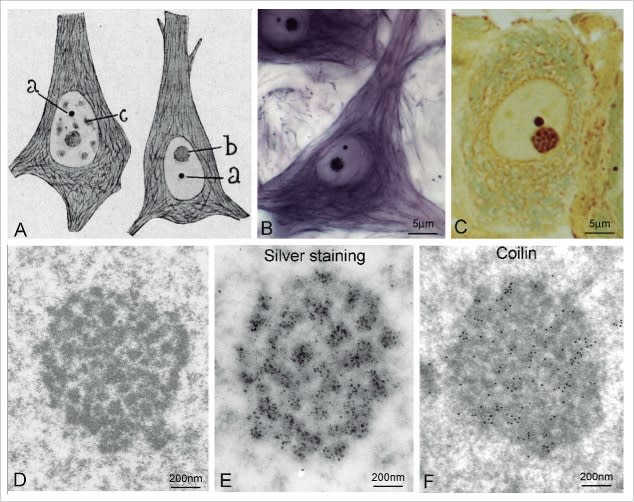
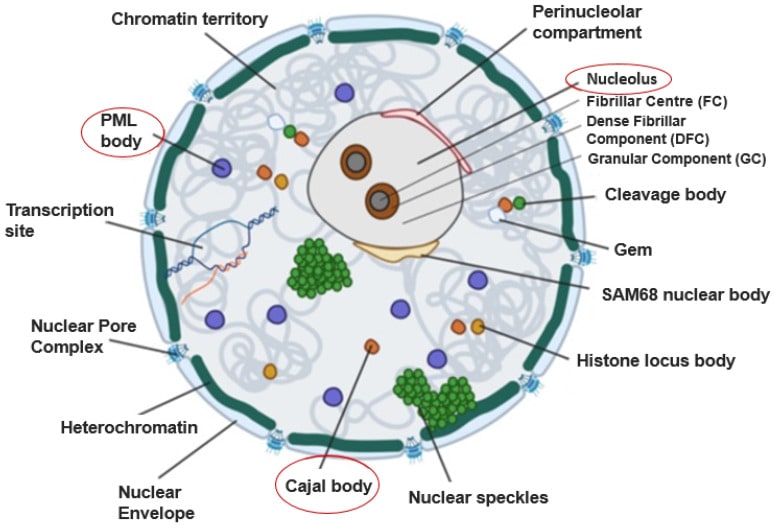
La estructura que Cajal describió como un apéndice del nucleolo ha sido radicalmente redefinida por la biología molecular. Hoy sabemos que el Cuerpo de Cajal (CB) no es un orgánulo estático, sino una arquitectura de la vida: una estructura dinámica y sin membrana, clasificada como un “condensado biomolecular”. Estos condensados son comparables a gotas efímeras que se autoorganizan dentro del nucleoplasma, formadas por un proceso físico conocido como separación de fases líquido-líquido. Su función primordial es concentrar proteínas y ARN específicos en un espacio infinitesimal, acelerando drásticamente la eficiencia de las reacciones bioquímicas sin la necesidad de una barrera física como una membrana.
Coilina: La Proteína Arquitecto
La identidad y la propia existencia del Cuerpo de Cajal dependen de una proteína clave: la p80-coilina. La coilina actúa como una proteína de andamiaje esencial; su capacidad para interactuar consigo misma y para unirse a una multitud de otras moléculas es lo que inicia la formación del condensado. Aunque está presente de forma difusa en el nucleoplasma, su enriquecimiento en estos cuerpos es lo que los define. La regulación de este proceso es de una complejidad exquisita, gestionada a través de modificaciones postraduccionales de la propia coilina —como la fosforilación, la metilación y la SUMOilación— que funcionan como interruptores moleculares para controlar sus interacciones, su estabilidad y, en última instancia, el ensamblaje y desensamblaje del Cuerpo de Cajal en sincronía con el ciclo celular. Por ejemplo, los CBs se desensamblan durante la mitosis y se vuelven a ensamblar al inicio de la interfase, un proceso regulado por la fosforilación de la coilina, que la protege de la degradación.
Un Crisol para la Información Genética
La función principal del Cuerpo de Cajal lo sitúa en el corazón mismo de la expresión génica. Es un centro neurálgico para la biogénesis, maduración y modificación de las ribonucleoproteínas nucleares pequeñas (snRNP), los componentes moleculares de la maquinaria de ayuste o splicing. El splicing es el proceso de edición crucial mediante el cual se eliminan las secuencias no codificantes (intrones) de los transcritos de ARN mensajero (ARNm) antes de que puedan ser traducidos a proteínas. El CB funciona como una «cadena de montaje» y un «centro de control de calidad» donde las snRNP son ensambladas, modificadas (mediante procesos como la 2’-O-metilación y la pseudouridilación guiadas por ARN pequeños específicos del CB o scaRNAs) y preparadas para su vital tarea.
La física de la separación de fases no es un mero detalle estructural; es la causa directa de la función del CB. Al crear un microambiente que concentra reactivos, el CB supera las limitaciones de la difusión en el nucleoplasma y acelera las reacciones bioquímicas, actuando como una fábrica de RNP de alta eficiencia. Esta eficiencia es precisamente lo que lo hace indispensable para la célula y, por tanto, un objetivo prioritario para los virus. La vulnerabilidad del CB a la manipulación viral no es una coincidencia, sino una consecuencia directa de su mecanismo operativo fundamental. Los virus han evolucionado para explotar el punto de mayor eficiencia de la célula, convirtiendo al CB en un nodo computacional dinámico que, si es controlado, permite dominar el flujo de la información genética celular.
El Archipiélago Nuclear: ¿Qué Vio Realmente Cajal?
La agudeza visual de Cajal, capaz de aislar el “cuerpo accesorio” del resto de estructuras, nos lleva a una pregunta fascinante: ¿fue también el primero en atisbar otros cuerpos nucleares, como los Cuerpos de PML, décadas antes de su descripción formal? Un análisis detallado de sus esquemas, como el que se muestra a continuación, nos ofrece una respuesta llena de matices. Recuerdos de mi vida > Sumario > Segunda parte, XXIV.



Este dibujo es la prueba irrefutable de que Cajal vio un universo de cuerpos nucleares, no solo uno. Con una precisión asombrosa, no solo etiqueta el nucleolo (‘a’) y su “cuerpo accesorio” (‘b’), sino que cataloga un ecosistema completo de estructuras: el “casquete cromático” (‘c’), el “grumo hialino” (‘e’), los “granitos basiófilos” (‘f’) y el “armazón fibrilar” (‘g’). Esto demuestra que su mirada penetró en la complejidad del núcleo a un nivel que nadie había alcanzado.
Es altamente probable que algunos de los diversos “granitos” y “grumos” que dibujó fueran, de hecho, lo que hoy conocemos como Cuerpos de PML. Sin embargo, no los agrupó bajo una única denominación ni los describió como una estructura singular y recurrente, a diferencia de como hizo de forma explícita con el “cuerpo accesorio”. La distinción que hoy hacemos entre un Cuerpo de Cajal, un Cuerpo de PML o un nuclear speckle es fundamentalmente molecular, definida por sus proteínas andamio (Coilina para el CB, proteína PML para el cuerpo de PML, etc.). El método de tinción de Cajal, basado en el nitrato de plata, era una técnica química que revelaba la morfología, no la composición proteica.
En resumen, este esquema no nos permite afirmar que Cajal “descubrió” los Cuerpos de PML, pero demuestra de una manera espectacular que fue el primero en cartografiar el archipiélago desconocido del núcleo celular. Vio las diferentes “islas”, aunque no tuviera las herramientas para dar un nombre geológico a cada una. Es, una vez más, una prueba de su genio visionario.
La Conexión Humana: SMN y la Atrofia Muscular Espinal (AME)
La importancia crítica del Cuerpo de Cajal para la salud humana queda dramáticamente ilustrada por su conexión con la Atrofia Muscular Espinal (AME), una devastadora enfermedad neurodegenerativa. La AME es causada por la deficiencia de la proteína Survival of Motor Neuron (SMN), un componente fundamental que se localiza prominentemente en los CBs. La proteína SMN es parte de un complejo esencial para el ensamblaje de las snRNP en el citoplasma. Una vez ensambladas, las snRNP, probablemente asociadas a SMN, son importadas al núcleo y dirigidas a los CBs para su maduración final.
La proteína SMN interactúa directamente con la coilina, y esta interacción es crucial para el correcto ensamblaje y la integridad del CB. En ausencia de coilina, ni SMN ni las snRNP se localizan correctamente en los CBs residuales. En pacientes con AME, la falta de SMN funcional conduce a la disrupción y pérdida de los CBs en las neuronas motoras, lo que resulta en un ensamblaje defectuoso de la maquinaria de splicing, parálisis progresiva y, en los casos más severos, la muerte. La pérdida de los Cuerpos de Cajal es, por tanto, un sello celular distintivo de la AME, estableciendo un vínculo inequívoco entre la integridad de este orgánulo y la supervivencia neuronal.
Ecos de un Pasado Olvidado: Cajal, el Microbiólogo
Aunque la historia recuerda a Cajal como el padre de la neurociencia, su carrera abarcó campos mucho más amplios, incluyendo una incursión decisiva en la microbiología y la salud pública. Esta faceta, a menudo olvidada, establece un poderoso paralelismo entre su lucha histórica contra los patógenos y la batalla celular que hoy se libra en el interior del Cuerpo de Cajal. En 1885, una devastadora epidemia de cólera asoló España, y Cajal, entonces catedrático en Valencia, se sumergió en el estudio de la enfermedad. Su trabajo no se limitó a la observación; desarrolló una innovadora «vacuna química» utilizando cultivos del bacilo del cólera inactivados por calor. Este enfoque, que introdujo el concepto de utilizar componentes microbianos muertos para generar inmunidad, fue pionero y se anticipó a trabajos similares y más reconocidos de investigadores estadounidenses.

Su compromiso con la lucha contra las enfermedades infecciosas trascendió el laboratorio. En el año 1900, Cajal fue nombrado primer director del Instituto de Sueroterapia, Vacunación y Bacteriología de Alfonso XIII, que más tarde se convertiría en el Instituto Nacional de Higiene. Aceptó el cargo por «responsabilidad patriótica», sacrificando su amada investigación personal para dotar a España de soberanía sanitaria en la producción de sueros y vacunas. Su gestión fue una batalla titánica contra la indiferencia política y la burocracia. La primera sede del instituto fue una antigua vaquería insalubre, y Cajal se vio consumido por lo que él mismo denominó la «tiranía del papel timbrado», llegando a adelantar dinero de su propio bolsillo para no detener la producción vital de vacunas.
Esta etapa de su vida establece una profunda resonancia temática con el tema central de este artículo. La lucha de Cajal para establecer una infraestructura de defensa sanitaria a nivel nacional es un reflejo, a escala social, de la lucha de la célula para montar una defensa antiviral a nivel subnuclear. El Instituto de Higiene fue la «piedra angular de la salud pública española»; el Cuerpo de Cajal es una piedra angular del procesamiento de ARN y, como veremos, de la defensa nuclear. La simetría es asombrosa: el Cajal histórico libró una guerra macroscópica contra una bacteria; la ciencia actual desvela una guerra microscópica contra los virus que tiene lugar en el mismo orgánulo que él descubrió.
El Campo de Batalla Nuclear: El Secuestro del Legado de Cajal
La importancia fundamental del Cuerpo de Cajal en el procesamiento del ARN no ha pasado desapercibida para los virus. A lo largo de la evolución, una amplia gama de virus, tanto de ADN como de ARN, han desarrollado estrategias convergentes para secuestrar esta estructura nuclear. Su objetivo es subvertir la arquitectura nuclear y la maquinaria de procesamiento de ARN del huésped para ponerla al servicio de su propia replicación. Los virus no se limitan a atacar el CB; lo remodelan estratégicamente, tratándolo como un recurso programable.
El hecho de que virus de familias tan diversas como Adenoviridae, Orthomyxoviridae y Flaviviridae converjan en el CB como objetivo es una prueba irrefutable de la centralidad de este orgánulo. No es un blanco casual, sino un “cuello de botella” fundamental en el control de la célula, un nexo donde los virus pueden priorizar sus propios ARN y desestabilizar los del huésped. Por ello, el estado del Cuerpo de Cajal puede considerarse un biomarcador de la infección viral, un “canario en la mina” nuclear.
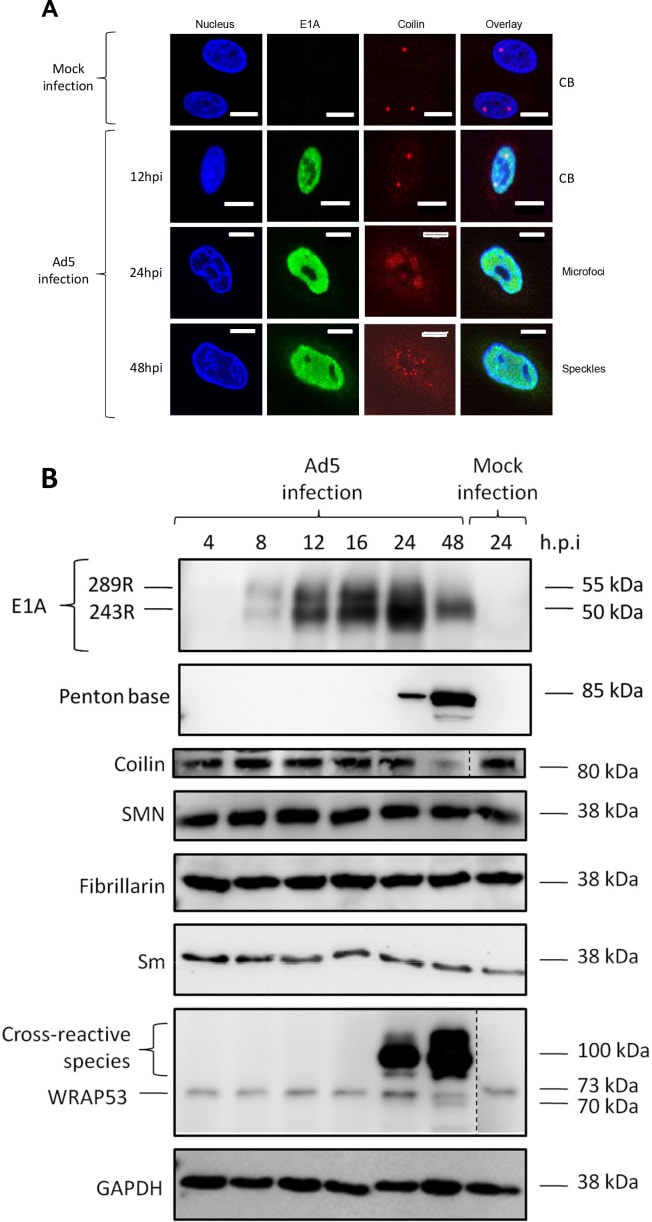
Tácticas de Secuestro Viral
Estas manipulaciones virales provocan cambios drásticos en la morfología, el número y la distribución de los Cuerpos de Cajal, visibles al microscopio. El arsenal de tácticas de secuestro es diverso y sofisticado:
-
Fragmentación: Virus como el de la gripe A o el Zika provocan que los CB se rompan en múltiples cuerpos más pequeños y numerosos, dispersando sus componentes.
-
Reorganización: El Adenovirus reorganiza los CB en estructuras únicas denominadas «microfocos» o «rosetas», que se posicionan estratégicamente junto a los centros de replicación del ADN viral.
-
Creación de Nuevos Cuerpos: Algunos virus, como el Virus Diminuto de Ratón (MVM), van un paso más allá y cooptan los componentes de los CB para construir fábricas de replicación viral completamente nuevas.
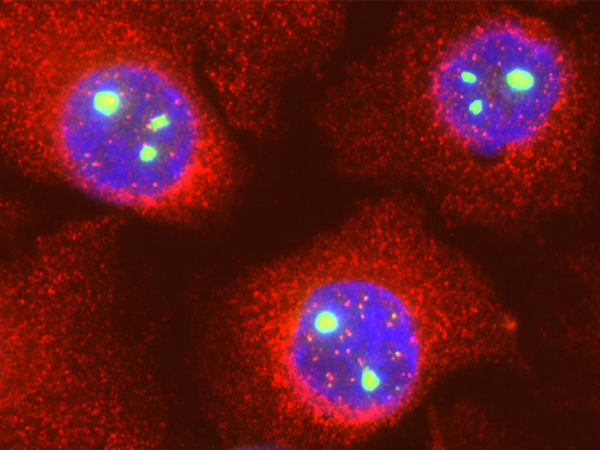
El caso del Adenovirus ofrece un ejemplo paradigmático de este golpe de estado molecular. La proteína viral L4-22K se une directamente y forma un complejo estable con la coilina del huésped. Este secuestro tiene un propósito muy específico: utilizar la coilina como un «salvoconducto» para facilitar la exportación de los ARN mensajeros virales desde el núcleo al citoplasma, un paso indispensable para la síntesis de nuevas proteínas virales. La depleción de coilina reduce drásticamente el rendimiento del virus al impedir la exportación de sus ARNm. En esencia, el virus convierte al arquitecto jefe del CB en un cómplice para ejecutar sus propios planos genéticos, como se ilustra en la Figura 6, que muestra la dramática reorganización de la coilina en microfocos tras la infección por Ad5.
La siguiente tabla resume la diversidad de estrategias que diferentes virus emplean para manipular el Cuerpo de Cajal, ilustrando la naturaleza central de este orgánulo en el conflicto virus-huésped.
**Familia Vírica/EjemploCambio Morfológico en los CBsMecanismo Molecular ClaveConsecuencia para la Infección ViralAdenovirus (Ad5)**Redistribución en «microfocos» o «rosetas» junto a los centros de replicación.La proteína viral L4-22K se une directamente a la coilina.Secuestro de la coilina para facilitar la exportación nuclear del ARNm viral.Virus de la Gripe AAumento del número, disminución del tamaño. Colocalización con la nucleoproteína viral (NP).Interacción con la NP viral.Posiblemente relacionado con el tráfico de componentes virales o la subversión de la maquinaria de splicing.**Virus del Zika (ZIKV)**Aumento del número y reducción del volumen.La proteína viral NS5 interactúa con la coilina, causando la disrupción del CB.Subversión de la arquitectura nuclear para favorecer la replicación viral.**Virus Diminuto de Ratón (MVM)**Reconstitución de los componentes del CB en nuevas fábricas virales (cuerpos APAR).Cooptación de componentes del CB, incluyendo la proteína SMN.Creación de centros de replicación viral dedicados y eficientes.**Virus de la Roseta del Cacahuete (GRV)**Reorganización en estructuras «similares a CB» que se fusionan con el nucleolo.La proteína viral ORF3 media la reorganización y fusión con el nucleolo.Secuestro de componentes celulares para el movimiento viral y la estabilización del ARN.
La Contraofensiva Celular: El Estado Antiviral y los Guardianes Mx
Frente a esta invasión, la célula no permanece pasiva. Ha desarrollado un sofisticado sistema de defensa de múltiples capas, y en el corazón de la respuesta innata se encuentra una familia de proteínas guardianas cuya historia se entrelaza con la lucha contra el mismo virus que manipula el Cuerpo de Cajal: la gripe.
La Señal de Alarma: El Sistema del Interferón
La primera línea de defensa de la célula es la detección del intruso. La presencia de patrones moleculares asociados a patógenos (PAMPs), como el ARN de doble cadena viral, activa sensores celulares que desencadenan una cascada de señalización. El resultado es la producción y secreción de citoquinas especializadas llamadas interferones (IFN) de tipo I (α/β) y tipo III (λ). Estas moléculas actúan como una sirena de alarma molecular. Se unen a receptores en la superficie de la célula infectada (acción autocrina) y de las células vecinas no infectadas (acción paracrina), induciendo la expresión de cientos de genes estimulados por interferón (ISG, por sus siglas en inglés). Este proceso establece un potente «estado antiviral» que prepara a las células para combatir la infección, creando un ambiente inhóspito para la replicación viral.
Los Guardianes del Genoma: Las Proteínas Mx (Mx1/MxA)
Entre los efectores más potentes y mejor estudiados de este arsenal de ISGs se encuentra la familia de proteínas de resistencia a mixovirus (Mx). Descubiertas hace más de 50 años gracias a la observación de que una cepa de ratones (A2G) era naturalmente resistente a la infección por el virus de la gripe, las proteínas Mx son GTPasas grandes pertenecientes a la superfamilia de la dinamina. Su expresión es estrictamente dependiente de los interferones de tipo I y III, y su función antiviral es autónoma: pueden proteger a las células de la infección incluso en ausencia de otros ISGs. Dos propiedades son absolutamente esenciales para su función: su capacidad para hidrolizar GTP (actividad GTPasa) y su habilidad para autoensamblarse formando oligómeros, como anillos y hélices.
Una Estrategia de Localización: La Defensa Nuclear vs. Citoplasmática
La eficacia de las proteínas Mx no reside únicamente en su bioquímica, sino en una tríada interdependiente de localización, oligomerización y actividad GTPasa. La disfunción en cualquiera de estos tres pilares anula su capacidad defensiva. La localización subcelular es un determinante clave de su espectro de acción. La proteína Mx1 de ratón, el prototipo de resistencia a la gripe, se acumula predominantemente en el núcleo. En cambio, su homóloga humana, la proteína MxA (codificada por el gen MX1), es casi exclusivamente citoplasmática. Esta divergencia evolutiva ha llevado a una especialización funcional: Mx1 es un guardián nuclear, idealmente posicionado para interceptar virus que, como la gripe, deben replicar su genoma en el núcleo. MxA, por otro lado, actúa como un centinela citoplasmático, interceptando componentes virales antes de que alcancen su destino nuclear, lo que le confiere un espectro antiviral más amplio contra virus que replican en distintos compartimentos celulares.
Característica Clave****Mx1 (Ratón)****MxA (Humana)****Especie PrototípicaRatón (cepa A2G)HumanoGenMx1MX1Localización CelularPredominantemente NuclearPredominantemente CitoplasmáticaEspectro Antiviral**Especialista, potente contra virus de replicación nuclear (ej. Gripe, Thogoto).Generalista, amplio espectro contra virus de ARN y algunos de ADN.Mecanismo vs. GripeBloquea la transcripción primaria del genoma viral dentro del núcleo.Intercepta los complejos de ribonucleoproteína (vRNP) virales en el citoplasma, impidiendo su importación nuclear o función posterior.Papel como BarreraModelo clave para estudiar la resistencia innata a la gripe en mamíferos.Actúa como una barrera entre especies contra la transmisión de virus zoonóticos (ej. gripe aviar).
El Mecanismo de Acción contra la Gripe: Desarmando el Complejo de Replicación Viral
El objetivo de las proteínas Mx en la infección por gripe es el corazón mismo del virus: el complejo de ribonucleoproteína viral (vRNP). Este complejo está formado por el genoma de ARN viral, que se encuentra recubierto por múltiples copias de la nucleoproteína (NP) y asociado a la polimerasa viral (compuesta por las subunidades PB1, PB2 y PA). Las proteínas Mx reconocen específicamente la NP dentro de este complejo. Tras el reconocimiento, las moléculas de Mx utilizan su capacidad de autoensamblaje para oligomerizarse y formar estructuras anulares que rodean y atrapan al vRNP. La hidrólisis de GTP probablemente impulsa cambios conformacionales que “aprietan la trampa”, desestabilizando el complejo viral e impidiendo su función. Específicamente, se ha demostrado que Mx1 interfiere directamente con la interacción crucial entre la NP y la subunidad de la polimerasa PB2, un paso indispensable para la transcripción y replicación del genoma viral.
La Carrera Armamentística Evolutiva
La proteína MxA humana no es solo un factor de restricción; es un guardián evolutivo que define, en parte, la barrera entre especies para los virus de la gripe. Es notablemente eficaz para restringir los virus de la gripe aviar, como el H5N1, lo que representa un obstáculo significativo para su transmisión a humanos. Sin embargo, los virus que han logrado establecerse en la población humana y causar pandemias, como la cepa de 1918 y la H1N1 de 2009, lo han hecho adquiriendo mutaciones adaptativas en su proteína NP. Estas mutaciones alteran la superficie de la NP en los sitios de reconocimiento de MxA, permitiendo al virus evadir esta primera línea de defensa innata. Por lo tanto, la vigilancia de las secuencias de NP de los virus de la gripe que circulan en reservorios animales, con un enfoque en las regiones que interactúan con MxA, se convierte en una herramienta de salud pública de importancia crítica para predecir el potencial pandémico de nuevas cepas.
Un Ecosistema de Defensa: La Arquitectura de la Inmunidad Nuclear
La respuesta antiviral no depende de efectores aislados como las proteínas Mx, sino de una red coordinada y una reorganización arquitectónica del núcleo. La señalización por interferón no solo induce la expresión de cientos de genes, sino que remodela el paisaje nuclear, fortaleciendo y conectando compartimentos funcionales para crear una red de defensa integrada y resiliente.
Los Cuerpos de PML como Centros de Mando Antivirales
Otro actor clave en esta defensa son los Cuerpos de la Leucemia Promielocítica (PML bodies), también conocidos como dominios nucleares 10 (ND10). Al igual que los CBs, son condensados biomoleculares sin membrana cuya formación y tamaño aumentan drásticamente en respuesta al interferón y a otros estreses celulares. Los cuerpos de PML actúan como “cuarteles generales” o centros de mando antivirales. Son sitios de intensa modificación postraduccional, especialmente la sumoilación, un proceso en el que se añade la pequeña proteína SUMO (Small Ubiquitin-like Modifier) a otras proteínas. Esta modificación puede alterar la localización, estabilidad o actividad de sus dianas, y los cuerpos de PML sirven como “fábricas de sumoilación” que activan y regulan numerosos factores de restricción viral y proteínas de respuesta al daño en el ADN.
La Colaboración entre Cuerpos Nucleares
Estos centros de mando no operan de forma aislada. Existe evidencia de una asociación física y funcional entre los Cuerpos de Cajal y los Cuerpos de PML. Se ha identificado una interacción directa entre la coilina, la proteína arquitecto del CB, y PIASy, un componente integral de los cuerpos de PML, que podría actuar como el puente molecular que une estos dos centros de defensa. Esta conexión sugiere un flujo de información o de componentes entre ellos. Además, la proteína Mx1 de ratón, el guardián nuclear contra la gripe, se ha encontrado asociada con componentes de los cuerpos de PML, lo que la sitúa físicamente en el corazón de la maquinaria de respuesta antiviral nuclear.
El Cuerpo de Cajal como Plataforma de ISGs
Esta red interconectada revela una nueva faceta del Cuerpo de Cajal. No es solo un objetivo pasivo para el secuestro viral, sino también una plataforma activa para la contraofensiva celular. En un estado de alerta inducido por el interferón, el CB recluta efectores de la inmunidad innata. Un ejemplo notable es la exonucleasa ISG20, otra proteína antiviral inducida por interferón, que se localiza predominantemente en los Cuerpos de Cajal. Este hallazgo consolida la visión del CB como un nodo integrador con una función dual. En tiempos de paz, es una fábrica para la biogénesis de RNP. En tiempos de guerra, se convierte en una plataforma militarizada que reúne componentes de defensa antiviral. La naturaleza del CB como un condensado líquido es clave para esta plasticidad funcional. Su composición puede cambiar dinámicamente: en ausencia de infección, está enriquecido con factores de procesamiento de ARN; tras la señal de interferón, puede reclutar nuevos componentes como los ISGs para cambiar su función. La célula no necesita construir una nueva fábrica; simplemente cambia el personal y las herramientas en la que ya existe.
El Cuerpo de Cajal como Sensor de Estrés Celular: Una Nueva Dimensión en la Inmunidad
La función del Cuerpo de Cajal trasciende la mera producción de componentes moleculares. Evidencias crecientes sugieren que tanto el CB como su proteína arquitecto, la coilina, actúan como sensores del estado de salud de la célula. Diversos tipos de estrés celular, como la radiación UV, el estrés osmótico o el daño en el ADN, provocan una rápida alteración en la estructura de los CBs y una redistribución de la coilina y otros de sus componentes. Esta respuesta no es un simple colapso, sino una reorganización activa que se cree que forma parte de una red de señalización de estrés más amplia, en la que el CB, en concierto con el nucleolo, juega un papel central.
La infección viral representa una de las formas más profundas de estrés celular. Desde esta perspectiva, la disrupción de los Cuerpos de Cajal por parte de los virus adquiere un nuevo significado. No se trata solo de secuestrar una fábrica, sino de desactivar un sistema de alarma. La coilina, en particular, ha demostrado estar implicada en la regulación de genes de la inmunidad y en la activación de mecanismos de defensa, especialmente en respuesta a infecciones virales. Se ha observado que la coilina es reclutada a sitios de daño en el ADN, un tipo de agresión que algunos virus provocan, y que su presencia modula la respuesta de reparación.
Por lo tanto, es plausible que la estrategia viral de fragmentar o reorganizar los CBs sea una táctica de contrainteligencia: al desmantelar estos centros de detección y señalización, el virus no solo se apropia de sus recursos, sino que también ciega a la célula ante su propia presencia, saboteando una respuesta inmune coordinada desde el mismo núcleo. Esta visión del Cuerpo de Cajal como un nexo de la inmunidad innata, un guardián que es neutralizado por el invasor, añade una capa de complejidad fascinante a la incesante batalla que se libra en el interior de nuestras células.
De la Observación a la Terapia: Nuevas Fronteras Antivirales
La misma característica que convierte al Cuerpo de Cajal en un objetivo principal para los virus —su papel central en el metabolismo del ARN— también lo convierte en una diana terapéutica excepcionalmente prometedora para el desarrollo de fármacos antivirales de amplio espectro. Si logramos comprender en detalle cómo los virus secuestran la coilina y desmantelan el CB, podremos diseñar intervenciones moleculares para bloquear este proceso y detener la infección.
Esta idea está dando lugar a un nuevo paradigma en el descubrimiento de fármacos: el desarrollo de «fármacos modificadores de condensados» (c-mods). A diferencia de los fármacos tradicionales, que suelen inhibir el sitio activo de una única enzima siguiendo un modelo de “llave-cerradura”, estas moléculas actuarían modulando las propiedades físicas del condensado en su conjunto. Podrían, por ejemplo, impedir su formación, forzar su disolución o alterar su composición para hacerlo inhóspito para el virus. La interacción específica entre una proteína viral y una proteína de andamiaje como la coilina representa un punto de intervención ideal.
La viabilidad de este enfoque no es meramente teórica. Una prueba de concepto contundente proviene del campo de las enfermedades neurodegenerativas, en particular de la Atrofia Muscular Espinal (AME). Esta devastadora enfermedad está causada por la deficiencia de la proteína SMN, un componente fundamental del Cuerpo de Cajal. La pérdida de los Cuerpos de Cajal es un sello celular distintivo de la AME, y su número se correlaciona inversamente con la gravedad de la enfermedad, lo que subraya la importancia crítica de la integridad del CB para la salud humana.
El extraordinario éxito de las terapias con oligonucleótidos antisentido (ASO) para la AME, como el Nusinersén, valida este enfoque. Estos fármacos actúan corrigiendo directamente el proceso de splicing del gen SMN2, lo que permite aumentar la producción de proteína SMN funcional y restaurar parcialmente la función del Cuerpo de Cajal. Este es un ejemplo de una terapia clínicamente exitosa que funciona manipulando la misma maquinaria de procesamiento de ARN que se ensambla y mantiene en el CB.
Existe una sorprendente convergencia terapéutica en el Cuerpo de Cajal: un virus lo secuestra para replicarse, mientras que una enfermedad genética surge de su disfunción. Una terapia exitosa para la AME funciona restaurándolo. Esto demuestra que la integridad funcional del CB es un pilar de la salud celular. El éxito clínico en AME proporciona una hoja de ruta directa para el desarrollo de antivirales dirigidos al huésped. Demuestra que las vías moleculares que convergen en el CB son “farmacológicamente accesibles” y que proteger su integridad es una estrategia terapéutica viable. Las lecciones aprendidas de la AME pueden informar directamente el diseño de una nueva generación de antivirales que busquen “blindar” al CB del ataque viral, ofreciendo potencialmente una protección de amplio espectro con una nueva generación de antivirales.
La Relevancia Perenne de un Sabio
La historia del Cuerpo de Cajal es un viaje extraordinario que cierra un círculo perfecto. El «cuerpo accesorio» que el ojo artístico de Cajal capturó por primera vez en 1903 es hoy un centro dinámico de la vida celular y un nexo de la inmunidad innata. La batalla que él libró contra el cólera en las calles de Valencia se libra ahora, a nivel molecular, contra una miríada de virus dentro de esa misma estructura que él descubrió.
El genio de Cajal no se limitó a la anatomía estática; fue un teórico de la función. Sus famosas flechas, dibujadas a mano para indicar la dirección del flujo de la información neuronal, fueron el germen de la teoría de circuitos en la que se basa toda la neurociencia. En su obra Textura del sistema nervioso, fue más allá, postulando que las células piramidales de la corteza cerebral eran las «células psíquicas», el sustrato material de las actividades más elevadas como el pensamiento y la memoria. Intuyó que la complejidad de estas células era la base de la inteligencia.
Existe una hermosa simetría conceptual entre los dos grandes legados de Cajal. Él conceptualizó el cerebro como una red de unidades discretas (neuronas) que procesan información. Más de un siglo después, su otro descubrimiento, el CB, se revela como un nodo dentro de una red de condensados nucleares que procesan la información genética. Hay una especie de fractalidad en su visión: los principios de organización y flujo de información que él postuló para el macrocosmos del cerebro se reflejan en el microcosmos del núcleo celular. La lucha por el control del Cuerpo de Cajal es, en esencia, una lucha por el control de la información de la célula. El gran tema unificador de la obra de Cajal, tanto en neurociencia como en biología celular, es la organización física del procesamiento de la información biológica.
El verdadero legado de Cajal no reside únicamente en sus descubrimientos, sino en su filosofía científica: una curiosidad incansable, una fe inquebrantable en la observación meticulosa y la profunda convicción de que la ciencia debe estar al servicio de la humanidad. Creía que la ciencia no era un coto reservado a genios, sino el resultado de la perseverancia y el «trabajo obstinado». Los investigadores que hoy desentrañan los secretos del Cuerpo de Cajal son los herederos directos de este espíritu. Su trabajo demuestra que, incluso en sus observaciones más aparentemente secundarias, la mirada de Cajal fue capaz de señalar el camino hacia el futuro. Como él mismo afirmó, la exploración del universo biológico es una empresa sin fin. La investigación sobre el Cuerpo de Cajal, un pequeño mundo dentro de la célula, es una prueba viviente de su visión.
«Mientras nuestro cerebro sea un misterio, el universo, reflejo de la estructura del cerebro, será también un misterio».
Santiago Ramón y Cajal
Rock & Roll



El profesor Angus Lamond y Cajal
El laboratorio Lamond, cuyo equipo puedes ver en la página web, es el creador del video “A Christmas Cajal”. En esta historia navideña, el momento científico clave es cuando logran grabar el movimiento del cuerpo de Cajal.
Bibliografía
-
Ramón y Cajal, S. (1903). Un sencillo método de coloración selectiva del retículo protoplasmático y sus efectos en los diversos órganos nerviosos. Trabajos del Laboratorio de Investigaciones Biológicas de la Universidad de Madrid, 2, 129–221.
-
Ramón y Cajal, S. (1899-1904). Textura del sistema nervioso del hombre y de los vertebrados. Madrid: Moya.
-
Ramón y Cajal, S. (1917). Recuerdos de mi vida: Historia de mi labor científica. Madrid: Moya.
-
Ramón y Cajal, S. (1885). Investigaciones experimentales sobre el microbio vírgula del cólera y sus inoculaciones profilácticas. Zaragoza: Tipografía del Hospicio Provincial.
-
Gall, J. G. (2000). Cajal’s «accessory body»: the first hundred years. Annual Review of Cell and Developmental Biology, 16, 273-300.
-
Gall, J. G. (2003). The centennial of the Cajal body. Nature Reviews Molecular Cell Biology, 4(12), 975-980.
-
Banani, S. F., Lee, H. O., Hyman, A. A., & Rosen, M. K. (2017). Biomolecular condensates: a new cellular organizing principle. Nature Reviews Molecular Cell Biology, 18(5), 285–298.
-
Lettin, L., Erbay, B., & Blair, G. E. (2023). Viruses and Cajal Bodies: A Critical Cellular Target in Virus Infection?. Viruses, 15(12), 2311.
-
Haller, O., & Kochs, G. (2020). Human MxA protein: A key player in innate immunity to influenza A virus. Journal of Experimental Medicine, 217(1), e20190933.
-
Bhat, P., Aksenova, V., Gazzara, M. R., et al. (2023). Influenza virus mRNAs encode determinants for nuclear export via the cellular TREX-2 complex. Nature Communications, 14(1), 2304.
-
White, L., Erbay, B., & Blair, G. E. (2023). The Cajal body protein p80-coilin forms a complex with the adenovirus L4-22K protein and facilitates the nuclear export of adenovirus mRNA. mBio, 14(5), e01459-23.
-
Finkel, R. S., Mercuri, E., Darras, B. T., et al. (2017). Nusinersen versus Sham Control in Infantile-Onset Spinal Muscular Atrophy. New England Journal of Medicine, 377(18), 1723–1732.
-
Arranz-Herrero, J., Izpura-Luis, S., Presa, J., et al. (2025). Swine influenza-modified pulmonary microbiota. Frontiers in Cellular and Infection Microbiology, 15, 1634469.
-
DeFelipe, J. (2006). Brain plasticity and mental processes: Cajal again. Nature Reviews Neuroscience, 7(10), 811-817.
Un sencillo método de coloración selectiva del retículo protoplasmáticoDescarga
Trabajos del Laboratorio de Investigaciones Biológicas de la Universidad de Madrid = Travaux du Laboratoire de Recherches Biologiques de l’Université de Madrid. SIMURG.
Trabajos del Laboratorio de Investigaciones Biológicas de la Universidad de Madrid = Travaux du Laboratoire de Recherches Biologiques de l’Université de Madrid. Vol. 2. 990001153000204201_V02.Descarga
Trabajos del Laboratorio de Investigaciones Biológicas de la Universidad de Madrid = Travaux du Laboratoire de Recherches Biologiques de l’Université de Madrid. SIMURG.


Comentarios
Para activar los comentarios: ve a giscus.app, introduce el repositorio
joseadserias-dotcom/cajal-digitaly reemplaza los IDs ensrc/layouts/Articulo.astro.